Version HTML de base


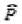
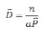
36
L’acoustique passive pourrait-elle être une alternative aux observations visuelles pour l’estimation des densités de populations ?
nous avons ainsi minimisé les fausses détections.
Deuxièmement, l’aire de détection couverte est déduite
d’une part de la distance maximale estimée par le modèle
RAM (différente en fonction du mois) et d’autre part en
tenant compte du masquage acoustique dû à la bathymé-
trie particulière autour de Crozet [20] (Fig. 2). Troisièmement,
l’histogramme des cris détectés en fonction des distances
va servir pour préciser la probabilité de détection .
Quatrièmement, il est nécessaire d’estimer le nombre de
cris émis par unité de temps pour cette espèce de céta-
cés. Ce dernier paramètre est particulièrement délicat à
établir et cela pour toutes les espèces de cétacés en géné-
ral. Marques et al. [10] propose de recourir à une balise
munie d’un hydrophone que l’on dépose sur le dos de l’ani-
mal et qui permet de l’enregistrer plusieurs heures pour
pouvoir faire une estimation robuste de
r
̂ . Il est cepen-
dant impossible de suivre cette approche dans notre cas.
Nous avons procédé autrement, en remarquant dans les
enregistrements, des séquences de cris (Fig. 5). Ces répé-
titions de sons sont interrompues par des périodes de
silence plus longues. Il est alors possible de caractériser
ces séquences et de définir un nombre de séquences par
unité de temps. Malheureusement, il y a plusieurs difficul-
tés à cette démarche. Tout d’abord, il n’est pas sûr que
ces séquences soient les mêmes pour tous les individus,
elles peuvent dépendre de l’âge, du comportement et
peut-être même du sexe. On n’est d’ailleurs actuellement
pas certains que les femelles baleines bleues émettent
ces sons [21]. Et des variations même faibles de ce para-
mètre entraînent des différences significatives dans les
estimations du nombre d’individus.
On peut s’affranchir de ces difficultés en proposant une
solution alternative consistant à considérer les données
comme une série temporelle. Tenant compte que d’une
part, un cétacé émet des sons de façon répétitive et que
d’autre part, certains individus seront silencieux pendant la
période d’enregistrement, nous avons introduit un proces-
sus de Poisson à 2 paramètres, l’un spatial et l’autre tempo-
rel. Après calculs de sa distribution et de sa fonction de
vraisemblance pour obtenir les estimations de ces 2 para-
mètres, on en déduit le nombre de cris par individu et par
conséquent la densité de population.
Conclusion
Connaître le nombre d’individus dans une population
de mammifères marins présents dans une zone est un
élément important pour faire du suivi de l’espèce, mais
également pour renseigner sur son environnement et sur
les possibles impacts des activités humaines. Il existe des
méthodes robustes basées sur les observations visuel-
les et l’enjeu actuel est de savoir s’il est possible de les
décliner aux observations acoustiques, qui peuvent être
soit complémentaires aux observations visuelles, soit les
seules possibles, comme ce fut le cas dans l’application
que nous avons détaillée. Toutefois, il reste encore quel-
ques difficultés, notamment l’estimation du nombre de cris
émis par unité de temps défini et par un individu.
Les perspectives seront donc soit de proposer des tech-
niques dédiées à certaines espèces, soit de proposer
des méthodes statistiques alternatives qui permettront,
à partir de simulations, de donner une estimation plus
fine de ce taux.
Remerciements
Ce travail a été financé notamment par l’association
Breach (www.breach-asso.org, France), l’association DIRAC
(France), le Commissariat à l’énergie atomique (www.cea.
fr) et le Ministère de l’écologie, de l’énergie, du dévelop-
pement durable et de la mer (http://www.developpement-
durable.gouv.fr).
Références bibliographiques
[1] IPCC. (2007). Climate Change 2007: The Physical Science Basis. Contribution
of Working Group I to the Fourth Assessment Report of the Intergovernmental
Panel on Climate Change. Cambridge University Press, Cambridge, UK and New
York, NY, USA.
[2] Simmonds M.P. and Eliott W.J. (2009). Climate change and cetaceans:
concerns and recent developments, J. of the Mar. Biol. Assoc. of the United
Kingdom, 89(1), 203–210
[3] Mellinger, D. K., Nieukirk, S. L., Matsumoto, H., Heimlich, S. L., Dziak, R. P.,
Haxel, J., Fowler, M., Meinig, C., and Miller, H.V. (2007). Seasonal occurrence of
North Atlantic right whale (Eubalaena glacialis) vocalizations at two sites on the
scotian shelf. Mar. Mamm. Sci. 23(4), 856–867
[4] Gandilhon, N., Adam, O., and Louis, M. (2010). Using passive acoustics for
marine mammal observations: multidisciplinary observatories, materials and
software, International Review of Physics, 4(1), p. 20-28 (A paraître)
[5] Seber, G. A. F. 1982. The estimation of animal abundance and related
parameters. 2nd edition, Griffin, London
[6] Hammond P. S. (1986). Estimating the size of naturally marked whale
populations using capture-recapture techniques, Rep. of the International Whaling
Commission, 8:252-282.
[7] Barlow, J., and Forney, K.A. (2007) Abundance and population density of
cetaceans in the California Current ecosystem, Fishery Bulletin, 105:509–526
[8] Buckland, S.T., Burnham, K. P., Anderson, D.R. and Laake, J.L. (1993).
Density estimation using Distance Sampling. London: Chapman and Hall.
[9] Buckland, S.T., Anderson, D.R., Burnham, K. P., Laake, J.L., Borchers, D.L.
and Thomas, L. (2004). Advanced Distance Sampling. Estimating abundance of
biological populations. Oxford University Press. Oxford, UK
[10] Marques, T.A., Thomas, L, Ward, J., DiMarzio, N. and Tyack, P.L. (2009),
Estimating cetacean population density using fixed passive acoustic sensors: An
example with Blainville’s beaked whales, J. Acoust. Soc. Am., 125(4): 1982-1994
[11] Clapham, PJ., Young, SB., and Brownell, JrRL. (1999). Baleen whales:
conservation issues and the status of the most endangered populations. Mammal
Rev, 29:35–60
[12] Ljungblad, D.K., Clark, C.W., Shimada, H. (1998). A comparison of sounds
attributed to pygmy blue whales (Balaenoptera musculus brevicauda) recorded
south of the Madagascar Plateau and those attributed to ‘true’ blue whales
(Balaenoptera musculus) recorded off Antarctica, Rep Int Whal Commiss.
48:439–442
[13] Rankin, S., Ljungblad, D., Clark, C., and Kato, H. (2005). Vocalisations
of Antarctic blue whales, Balaenoptera musculus intermedia, recorded during
the 2001/2002 and 2002/2003 IWC/SOWER circumpolar cruises, Area V,
Antarctica. J Cetacean Res Mngmt. 7:13–20
[14] Stafford, K.M., Mellinger, D.K., Moore, S.E., and Fox, C.G. (2007). Seasonal
variability and detection range modeling of baleen whale calls in the Gulf of
Alaska, 1999–2002. J Acoust Soc Am. 122:3378–3390
[15] Samaran, F., Adam, O., Motsch, J.-F., and Guinet C. (2008). Definition of the
Antarctic and Pygmy Blue whale call templates. Application to fast automatic
detection. Can Acoust. 36:93-102
[16] Samaran, F., Adam, O., Guinet, C., Motsch, JF. And Cansi, Y. (à paraître).
Source level estimation of two blue whale subspecies in Southwestern Indian
Ocean, J. Acoust. Soc. Am.
[17] Samaran, F. (2008). Analyse des signaux acoustiques d’origine biologique
enregistrés dans l’Océan Indien : Implication dans le recensement et le suivi
des mouvements saisonniers des cétacés. PhD thesis, University of La Rochelle,
France
[18] Collins, MD (1993). A split-step Padé solution for the parabolic equation
method. J. Acoust. Soc. Am. 93:1736–1742 (http://oalib.hlsresearch.com).
[19] Samaran, F., Adam, O., and Guinet, C. (à paraître). Detection range
modelling of blue whale calls in the Southwestern Indian Ocean. Applied
Acoustics.
[20] Samaran, F., Adam, O., and Guinet, C. (à paraître). Discovery of a mid-ocean
sympatric of southern blue whale subspecies. Endangered species research
[21] Oleson EM, Calambokidis J, Burgess WC, McDonald MA, LeDuc CA,
Hildebrand JA. (2007). Behavioral context of call production by eastern North
Pacific blue whales. Mar Ecol Prog Ser. 330:269–284
[22] McDonald, Hildebrand and Mesnick (2009). Worldwide decline in tonal
frequencies of blue whale songs, Endangered species research, 9:13-21